Hidrofobik mos kelmaslik - Hydrophobic mismatch
Hidrofobik mos kelmaslik ning qalinliklari orasidagi farq hidrofob a mintaqalari transmembran oqsili va biologik membrana u qamrab oladi.[1] Hidrofob yuzalarni suvga noxush ta'sirlanishiga yo'l qo'ymaslik uchun transmembran oqsillarining gidrofobik mintaqalari atrofdagi gidrofobik (lipidatsil zanjir) mintaqaning qalinligi taxminan bir xil bo'lishi kutilmoqda. lipidli ikki qatlam. Shunga qaramay, bir xil membrana oqsilini har xil qalinlikdagi ikki qavatli qatlamlarda uchratish mumkin.[2] Eukaryotik hujayralarda plazma membranasi endoplazmatik retikulum membranalariga nisbatan qalinroq bo'ladi. Shunga qaramay, plazma membranasida mavjud bo'lgan barcha oqsillar dastlab ribosomalarda sintez qilinganda endoplazmatik retikulumga qo'shiladi.[3] Transmembranli peptidlar yoki oqsillar va ularning atrofidagi lipidlar turli xil usullar bilan hidrofobik mos kelmasligi uchun moslasha oladi.
Mos kelmaslik uchun mumkin bo'lgan moslashuvlar
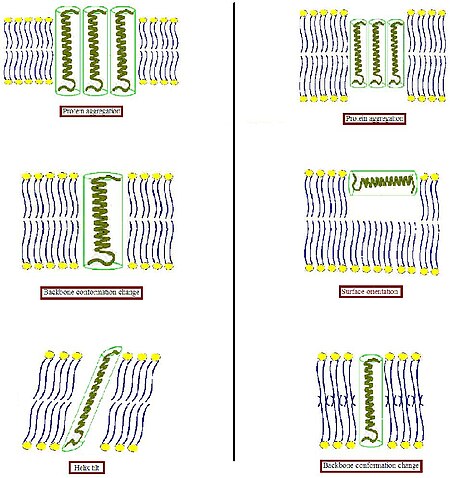
Hidrofobik sirtlarni hidrofil muhitga salbiy ta'sirini oldini olish uchun biologik membrana bunday nomuvofiqlikka biroz moslashishga intiladi. Boshqa turli xil tizimlarda, ajralmas oqsil hajmi va shakli mos lipidlar bilan o'zini o'rab olishga intilishidir.[4] Oqsillar nisbatan qattiq bo'lganligi sababli, lipidli uglevodorod zanjirlari egiluvchanligi sababli, hidrofobik mos kelish holatini lipid zanjirlarini cho'zish, siqish va / yoki egish orqali bajarish mumkin.[5][6][7]
- Transmembran oqsilining gidrofob qismi gidrofobik ikki qavatli qalinlikka (rasmning chap qismi) to'g'ri kelmaydigan darajada qalin bo'lsa, oqsil ta'sir qiladigan hidrofobik maydonni minimallashtirish yoki ularning samarali gidrofobik qalinligini kamaytirish uchun moyillikni kamaytirish uchun membranada to'planishi mumkin. Shuningdek, ular interfeys yaqinidagi hidrofob va gidrofil yon zanjirlarning yo'nalishini o'zgartirib, boshqa konformatsiyani qabul qilishlari mumkin. Lipidlar o'z navbatida o'zlarining asil zanjirlarini cho'zish orqali membrana qalinligini modulyatsiya qilishi yoki hattoki boshqa agregat turiga qo'shilishi va shu bilan ikki qavatli tashkilotni buzishi mumkin.
- Transmembran oqsilining hidrofob qismi hidrofobik ikki qavatli qalinlikka (rasmning o'ng qismida) to'g'ri kelmaydigan darajada kichik bo'lsa, bu yana oqsillarni to'planishiga yoki magistral konformatsiyasi va / yoki yon zanjir yo'nalishidagi o'zgarishlarga olib kelishi mumkin. Juda qisqa peptidlar qo'shimcha ravishda sirt lokalizatsiyasini o'z ichiga olmaydi. Lipidlar o'zlarining asil zanjirlarini tartibsizlantirish orqali samarali ikki qavatli qalinlikni kamaytirishi yoki teskari naychali bo'lmagan strukturani hosil qilish uchun ikki qavatli tashkilotni buzishi mumkin. Ushbu mumkin bo'lgan moslashuv usullarining kombinatsiyasi ham paydo bo'lishi mumkin.
Oqsillarni birlashishi
Mouritsen va Bloom beri[8] lipidlarning moslashuvi va ularning "matras modeli" ning o'ta mos kelmasligi paytida oqsillarni segregatsiyasini induktsiyalashni o'z ichiga olgan batafsil termodinamik modelni taklif qildi, nomuvofiqlikni keltirib chiqaradigan oqsillarni birlashishi to'g'risida qo'shimcha tushuncha olindi.[9] Shuningdek, hidrofobik mos kelmaslik suyuqlikning ikki qatlamida oqsillarni birlashishiga olib kelishi mumkinligi to'g'risida ba'zi eksperimental dalillar asoslandi.[10][11][12] Turli zanjir uzunligi bilan to'yingan va to'yinmagan suyuq PC ikki qavatli qatlamlarida qayta tiklangan bakteriorhodopsin bo'yicha elektron mikroskopik tadqiqotlar shuni ko'rsatdiki, oqsillar birlashishi faqat juda katta nomuvofiqlik bilan sodir bo'lgan va ikki qavatli qalinliklar 4 angstrom qalinroq va 10 angstrom ingichka hidrofobik qalinligidan ingichka. muhim birlashma induksiyasiz oqsilga ruxsat beriladi.[13]
Spiral nishab
Agar peptid yoki oqsilning hidrofob qismi membranani qoplash uchun juda uzun bo'lsa, moyillik ham mumkin. E. coli laktoza permeazasi bo'yicha avvalgi tadqiqotlar shuni ko'rsatdiki, PE / PG (3/1) lipid ikki qavatli qatlamida oqsil tiklanganda, tarkibida protein miqdori ko'payganda spiral moyilligi ko'payadi.[14] Ushbu nishab lipidlar tartibining pasayishi bilan birga kechdi, bu esa ikki qavatli qalinlikning pasayishiga olib keladi, bu esa mos kelmaslik bilan bog'liq javob ekanligini anglatadi. Membranani bir necha marta qamrab oladigan katta oqsillarda spiral moyillikning o'zgarishi lipidlar paketiga unchalik ta'sir qilmasdan sodir bo'lishi mumkin. Shu bilan birga, bitta transmembranli spiral uchun, burilish atrofdagi lipidlarga ikki qavatli spiralni joylashtirish uchun kuchlanishni keltirib chiqarishi mumkin. Shunday qilib, katta darajadagi burilish bitta transmembran oqsillari uchun unchalik qulay bo'lmagan variant bo'lishi mumkin.
Yuzaki yo'nalish
Nisbatan kichik gidrofobik peptidlar membranaga qo'shilmasligi mumkin va bunga javoban membrana yuzasida yo'nalish mavjud. Eksperimental dalillar sun'iy peptidga asosan leykinlarning 19 ta aminokislota gidrofobik ketma-ketligi va ikkala tomoniga ankraj qoldiqlari sifatida lizinlar bilan yonboshlab, sun'iy peptid ustida flüoresan tadqiqotida ko'rsatildi.[15] Natijalar shuni ko'rsatdiki, dominant transmembranadan peptidning parallel yo'nalishiga o'tish xolesterin qo'shilishi yoki lipid zanjiri uzunligini oshirish orqali ikki qavatli qalinlikni modulyatsiya qilish orqali yuzaga kelishi mumkin.
Magistral konformatsiyani o'zgartirish
Lipidli ikki qatlamli peptidlar va oqsillarning konformatsiyasi uchun mos kelmaslik oqibatlari to'g'risida batafsil ma'lumot olish uchun eng kichik membranani qamrab oluvchi peptidlar eng mos keladi. Hali ham ba'zi tadqiqotlar kerak.
Mos kelmaslik effektlari uchun nazariyalar
Mos kelmaslikning energiya xarajatlari va termodinamik ta'sirini tavsiflash uchun turli xil nazariy yondashuvlar qo'llanilgan, shu jumladan membranani elastik qatlam sifatida davolash[16][17]yoki mikroskopik usul.
To'shak modeli

To'shak modeli 1984 yilda Mouritsen va Blyum tomonidan fenomenologik nazariya yondashuvi sifatida taklif qilingan.[18] Bu nodavlat eritmalar nazariyasiga asoslangan ikki komponentli haqiqiy echim nazariyasidir va shu sababli fazalarni ajratishga imkon beradi. O'zlarining modellarida ular mos kelmaslik natijasida yuzaga kelgan membrana sirtining to'lqinlarida to'plangan energiyani lipidlar va oqsillarning elastik xususiyatlariga bog'laydi. Ular lipidlarning mikroskopik detallarini o'z ichiga olmaydi, ammo sof lipidlar tizimining ma'lum termodinamik xususiyatlaridan foydalanadi. Ular tarkibiga mos kelmaslik natijasida kelib chiqqan bilvosita lipid-oqsil o'zaro ta'sirlari, shuningdek lipid ikki qatlamining gidrofobik qismlari va oqsillar o'rtasidagi to'g'ridan-to'g'ri lipid-oqsil van der Vaalsga o'xshash o'zaro ta'sirlar kiradi. Lipid-oqsilli hidrofobik mos kelmaslik bilan bog'liq ortiqcha "hidrofob ta'sir" va oqsil yaqinidagi lipid zanjirlarining elastik deformatsiyaning erkin energiyasi. O'zaro ta'sir potentsiallari membrana xususiyatlarini termodinamik va mexanik o'lchovlardan olingan eksperimental ma'lumotlar asosida baholanadi.
Monte-Karlo simulyatsiyasi sxemasi
To'shak modeli keyinchalik Sperotto va Mouritsen tomonidan Monte-Karlo simulyatsiyasi sxemasida takrorlangan.[19] Ular Pinkning 10-davlat modeli bo'yicha tasniflangan lipidlarning turli xil mikrostatlariga imkon berishdi.[20] shuning uchun toza lipidli ikki qatlamli faza o'tishini ta'minlash. Modelning ushbu versiyasi tizimning mikroskopik xususiyatlari va uning termodinamik harakati o'rtasida bog'liqlikni ta'minlaydi.
Molekulyar nazariya
Membrananing lipid zanjirlarining molekulyar nazariyasida peptidlar, ularning hidrofob uzunligi bilan, lipid zanjirlari konfiguratsiyasida chegara shartini ta'minlovchi sifatida ko'rib chiqilgan.[21] Molekulyar modellashtirish lipid bosh guruhining surilishi va membrana erituvchisi sirt tarangligini tavsiflovchi fenomenologik erkin energiya qo'shimchalari bilan birlashtirildi. Dyuk va boshq.[22].
Hidrofobik mos kelmaslik va spiralning qiyshayishini eksperimental tadqiqotlar
Membrana oqsillarining nomuvofiqlikka javobini bilish turli xil eksperimental tadqiqotlar natijasida olingan. Turli xil eksperimental yondashuvlar yuqorida aytib o'tilgan gipotetik molekulyar reaktsiyalarning hissalari haqida har xil tushuncha beradi. Masalan, lyuminestsent yoki paramagnitik yorliqli guruhlar bilan jihozlangan oqsillar yoki peptidlar floresan spektroskopiyasida va elektron spin rezonansi tadqiqotlar. Bular oqsil-lipid o'zaro ta'sirining ham, oqsil-oqsilning o'zaro ta'sirining ham (birlashma uslubidagi reaktsiyaga xos) molekulyar tafsilotlarini va ularning (noto'g'ri) kelishuv sharoitlariga qanday ta'sir qilishini aniqlashi mumkin. Membrana qalinligi funktsiyasi sifatida spiralning burilishini o'rganish ham foydalandi qattiq holatdagi NMR texnikalar, xususan spiral burilish burchagi to'g'risida to'g'ridan-to'g'ri tushunishni ta'minlaydigan yo'naltirilgan membranalar yordamida. Model membranani qamrab oluvchi peptidlarni (masalan, WALP peptidi ) reaktsiyaga ta'sir qiluvchi turli xil omillar, jumladan membrana tarkibi, peptidlar ketma-ketligi va xususan, interfeyslararo ankraj qoldiqlari borligi to'g'risida tushuncha berdi. So'nggi yillarda katta yutuqlar Rentgenologik kristallografiya va elektron mikroskopi texnikasi katta oqsillarning lipidlar bilan o'zaro ta'siri haqida yangi tushunchalar berdi. Bunga kristallangan kaltsiy nasosi oqsilida spiralni burish haqidagi tushunchalar misol bo'la oladi [23].
Mos kelmaslikning biologik ahamiyati
Hidrofobik mos kelmaslik oqsillarni saralash va shakllanishi lipidli raftlar.
Oqsillarni saralash
Eukaryotik hujayralarda xolesterin darajasi sekretsiya yo'li orqali ortadi, endoplazmatik to'rdan Goljiga plazma membranasiga,[24] membrana qalinligining bir vaqtda o'sishini taklif qiladi. Bunga muvofiq, bitta spanli plazma membranasi oqsillarining transmembran segmentining o'rtacha uzunligi, odatda Golgi oqsillarining o'rtacha uzunligidan besh aminokislotaga ko'p.[25] Goljidagi oqsillarni saralash shu uzunlik farqiga asoslangan bo'lishi mumkinligi to'g'risida eksperimental dalillar olingan: odatda Goljida yashovchi bir nechta oqsillar uchun ularning hidrofob uzunligini oshirish oqsillarni plazma membranasiga yo'naltirishi yoki aksincha, plazma membranasidan oqsillarning gidrofob uzunligini pasayishi ularning Goljida tutilishiga olib kelishi mumkin.[26][27]
Lipit sallar
Raftlar - bu xolesterin, sfingomielin (SM) va ba'zi membrana oqsillari bilan boyitilgan membrana domenlari.[28][29] Raftlar signallarni o'tkazish, endotsitoz, apoptoz, oqsil savdosi va lipidlarni boshqarish kabi ko'plab fiziologik jarayonlarda taxminiy rollarga ega.[30][31][32][33][34] Raft lipidlari odatda to'yingan uglevodorod zanjirlariga ega.[35] Lipidli raftorlar lipidning ikki qavatli qatlamiga nisbatan yuqori hidrofobik qalinlikka ega, bu esa ko'proq gidrofobik qalinligi bo'lgan transmembran oqsillarini lipidli raftorlarga imtiyozli ravishda ajratilishiga olib kelishi mumkin.[36][37]
Shuningdek qarang
Adabiyotlar
- ^ Dyuk, D .; Li, X. J.; Katsov, K .; Schick, M. J. Chem. Fizika. 2002, 116, 10478-10484.
- ^ Killian, J.A. Biochimica et Biofhysica Acta. 1998, 1376, 401-416.
- ^ van Meer, G. Annu. Rev. Cell Biol. 1989, 5, 247-275.
- ^ Fattal, D. R .; Ben-Shoul, A. Biofiz. J. 1993, 65, 1795-1809.
- ^ Nezil, F. A .; Bloom, M. Biofhys. J. 1992, 61, 1176-1183.
- ^ Kurrle, A .; Riber, P .; Sackmann, E. Biokimyo. 1990, 29, 8274-8282.
- ^ Chjan, Y .; Lyuis, A. H.; Xodjes, R. S .; McElhaney, R. N. Biokimyo. 1992, 31, 11579-11588.
- ^ Mouritsen, O.G .; Bloom, M. Biofhys. J. 1984, 46, 141-153.
- ^ Mouritsen, O.G .; Biokimyo. Biofiz. Acta. 1998, 1376, 245-266,
- ^ Lyuis, B.A .; Engelman, D.M. J. Mol. Biol. 1983, 166, 203-210.
- ^ Ryba, N.J .; Marsh, D. Biokimyo 1992, 31, 7511-7518.
- ^ Kusumi, A .; Hyde, J.S. Biokimyo 1982, 21, 5978-5983.
- ^ Lyuis, B.A .; Engelman, D.M. J. Mol. Biol. 1983, 166, 203-210.
- ^ Le Coutre, J .; Narasimxon, L.R. ; Kumar, C .; Patel, N .; Kaback, HR Proc. Natl. Akad. Ilmiy ish. AQSh 1997, 94, 10167-10171.
- ^ Ren, J .; Lew, S .; Vang, Z.; London, E. Biokimyo 1997, 36, 10213-10220.
- ^ Dyuk, D .; Li, X. J .; Katsov, K .; Schick, M. J. Chem. Fizika. 2002, 116, 10478-10484.
- ^ Fattal, D. R .; Ben-Shoul, A. Biofiz. J. 1993, 65, 1795-1809.
- ^ Mouritsen, O.G .; Bloom, M. Biofhys. J. 1984, 46, 141-153.
- ^ Sperotto, M. M.; Mouritsen, O. G. Eur. Biofiz. J. 1991, 19, 157-168.
- ^ Pushti, D. A .; Chapman, D. Prok. Natl. Akad. Ilmiy ish. AQSH. 1979, 76, 1542-1546.
- ^ Fattal D. R.; Ben-Shoul, A. Biofiz. J. 1993, 65, 1795-1809
- ^ Dyuk, D .; Li, X. J.; Katsov, K .; Schick, M. J. Chem. Fizika. 2002, 116, 10478-10484.
- ^ Norimatsu, Y., Xasegava, K., Shimizu, N., Toyoshima, C. Tabiat 2017, 545, 193-198
- ^ Silvius, J.R .; McElhaney, R.N. Mumkin. J. Biokimyo. 1978, 56, 462-469.
- ^ Bretscher, M.S .; Munro, S. Fan 1993, 261, 1280-1281.
- ^ Munro, S. EMBO J. 1995, 14, 4659-4704.
- ^ Masibay, A.S .; Balaji, P.V .; Boeggeman, E.E .; Qasba, P.K. J. Biol. Kimyoviy. 1993, 268, 9908-9916.
- ^ Simons, K .; Tomre, D. Nat. Rev. Mol. Hujayra biol. 2000, 1, 31-39.
- ^ Simons, K .; Ikonen, E. Tabiat. 1997, 387, 569-572.
- ^ Simons, K .; Ikonen, E. Tabiat. 1997, 387, 569-572.
- ^ Tomas, J. L .; Xoloka, D .; Berd, B .; Veb, W. W. J. Cell Biol. 1994, 125, 795-802.
- ^ De Yong, K .; Geldverth, D .; Kuypers, F. A. Biokimyo. 1997, 36, 6768–6776.
- ^ Edidin, M. Trends Cell Biol. 2001, 11, 492-496.
- ^ Anderson, R. G. V.; Jacobson, K. Science. 2002, 296, 1821-1825.
- ^ Ohvo-Rekila, H.; Ramstedt, B .; Leppimaki, P.; Slotte, J. P. Prog. Lipid rez. 2002, 41, 66-97.]
- ^ Van Meer, G. Fan. 2002, 296, 855-856.
- ^ McIntosh, T. J. Chem. Fizika. Lipidlar. 2004, 130, 83-98.