Inson evolyutsiyasining xronologiyasi - Timeline of human evolution

The inson evolyutsiyasining xronologiyasi evolyutsion nasabidagi asosiy voqealarni bayon qiladi zamonaviy inson turlari, Homo sapiens davomida hayot tarixi, taxminan 4,2 milliard yil oldin boshlangan so'nggi evolyutsiya ichida H. sapiens paytida va undan keyin Oxirgi muzlik davri.
Bu turli xil qisqacha tushuntirishlarni o'z ichiga oladi taksonomik darajalar inson nasabida. Vaqt chizig'i asosiy qarashlarni aks ettiradi zamonaviy taksonomiya printsipiga asoslanib filogenetik nomenklatura; ochiq konsensusga ega bo'lmagan ochiq savollar bo'lsa, asosiy raqobat imkoniyatlari qisqacha bayon etilgan.
Taksonomik darajalarga umumiy nuqtai
Jadvalining umumiy ko'rinishi taksonomik reyting ning Homo sapiens (har bir daraja uchun yosh ko'rsatkichlari bilan) quyida ko'rsatilgan.
| Rank | Ism | Umumiy ism | Million yillar oldin (boshlanish) | |
|---|---|---|---|---|
| Hayot | 4,200 | |||
| Arxeya | ||||
| Domen | Eukaryota | Eukaryotlar (shilimshiq qoliplari va tegishli) | 2,100 | |
| Podiata | ||||
| Unikonts | ||||
| Obazoa | ||||
| Opisthokonts | Xolozoa + Qo'ziqorinlar s.l. | 1,300 | ||
| Xolozoa | 1,100 | |||
| Filozoa | Choanozoa + Filastereya | |||
| Choanozoa | Choanoflagelatlar + Hayvonlar | 900 | ||
| Qirollik | Animalia | Hayvonlar | 610 | |
| Subkingdom | Eumetazoa | |||
| Paraxoksozoa | ||||
| Bilateriya | Triploblastlar / qurtlar | 560 | ||
| Nefrozoa | ||||
| Deuterstomalar | ||||
| Filum | Chordata | Xordatlar (umurtqali hayvonlar va bir-biriga yaqin umurtqasizlar) | 530 | |
| Olfaktorlar | ||||
| Subfilum | Omurgalar | Baliq / umurtqali hayvonlar | 505 | |
| Infrafilum | Gnathostomata | Jag'li baliq | 460 | |
| Teleostomi | Suyakli baliq | 420 | ||
| Sarcopterygii | Lob qanotli baliq | |||
| Superklass | Tetrapoda | Tetrapodlar (to'rt oyoqli hayvonlar) | 395 | |
| Amniota | Amniotlar (tuxumlari to'liq quruqlikdagi tetrapodlar "amnios bilan jihozlangan" ) | 340 | ||
| Sinapsida | Proto-sutemizuvchilar | 308 | ||
| Terapsid | Tananing ostidagi a'zolar va boshqa sutemizuvchilarning xususiyatlari | 280 | ||
| Sinf | Sutemizuvchilar | Sutemizuvchilar | 220 | |
| Subklass | Theria | Tirik yoshni tug'diradigan sutemizuvchilar (ya'ni tuxum qo'ymaydigan) | 160 | |
| Infraklass | Evteriya | Plasental sutemizuvchilar (ya'ni, marsupials) | 125 | |
| Kattaroq | Boreoutheria | Supraprimates, (aksariyat) tuyoqli sutemizuvchilar, (eng) yirtqich sutemizuvchilar, kitlar va yarasalar | 124–101 | |
| Superorder | Evarxontoglires | Supraprimatlar: primatlar, kolugolar, daraxtlar, kemiruvchilar va quyonlar | 100 | |
| Katta tartib | Euarchonta | Primatlar, kolugos va daraxt daraxtlari | 99–80 | |
| Mirorder | Primatomorfa | Primatlar va kolugolar | 79.6 | |
| Buyurtma | Primatlar | Primatlar / Plesiadapiformes | 75 | |
| Suborder | Xaplorrin | "Quruq burunli" (so'zma-so'z "oddiy burunli") primatlar: buzadigan amallar va maymunlar (shu jumladan) maymunlar ) | 63 | |
| Infraorder | Simiiformes | maymunlar (maymunlar bilan birga) | 40 | |
| Parvorder | Katarrin | "Burun pastga" primatlar: maymunlar va keksa dunyo maymunlari | 30 | |
| Superfamily | Gominoideya | Maymunlar: buyuk maymunlar va maymunlar (gibbonlar) | 28 | |
| Oila | Hominidae | Ajoyib maymunlar: odamlar, shimpanzeler, gorillalar va orangutanlar hominidlar | 20–15 | |
| Subfamily | Gomininlar | Odamlar, shimpanzeler va gorillalar (Afrika maymunlari)[1] | 14–12 | |
| Qabila | Gominini | Ikkalasini ham o'z ichiga oladi Homo, Pan (shimpanze), lekin emas Gorilla. | 10–8 | |
| Obuna bo'lish | Gominina | Jins Homo va yaqin odam qarindoshlari va ajdodlari bo'linish dan Pan- bu homininlar | 8–4[2] | |
| (Tur) | Ardipitek s.l. | 6-4 | ||
| (Tur) | Avstralopitek | 3 | ||
| Jins | Homo (H. Habilis) | Odamlar | 2.5 | |
| (Turlar) | H. Erectus s.l. | |||
| (Turlar) | H. heidelbergensis s.l. | |||
| Turlar | Homo sapiens | Anatomik jihatdan zamonaviy odamlar | 0.8–0.3[3] |
Xronologiya
Bir hujayrali hayot
| Sana | Tadbir |
|---|---|
| 4.1 Ga (milliard.) yil avval) | Eng qadimgi hayot paydo bo'ladi. |
| 3.9 Ga | Hujayralar o'xshash prokaryotlar paydo bo'ladi. |
| 3,5 Ga | Bu kislorodli fotosintezning birinchi ko'rinishini va shuning uchun ko'p miqdordagi atmosferaning paydo bo'lishini anglatadi kislorod Yerda. |
| 2,5 Ga | Kisloroddan foydalanadigan birinchi organizmlar. 2400 yilgacha, deb ataladi Ajoyib oksigenatsiya hodisasi, hayotning kislorodgacha bo'lgan anaerob shakllari kislorod ishlab chiqaruvchilari tomonidan yo'q qilindi. |
| 2,1 ga | Keyinchalik murakkab hujayralar paydo bo'ladi: the eukaryotlar. |
| 1,2 Ga | Jinsiy ko'payish rivojlanadi, tezroq evolyutsiyaga olib keladi[4] bu erda genlar har bir avlodda aralashib, keyingi tanlov uchun katta o'zgarishlarga imkon beradi. |
| 0,9 Ga | The xanoflagellatlar ga o'xshash ko'rinishi mumkin ajdodlar butun hayvon qirollik va xususan, ular to'g'ridan-to'g'ri ajdodlari bo'lishi mumkin gubkalar.[5][6] Proterospongiya (Choanoflagellata a'zolari) - bu barcha hayvonlarning ajdodlari qanday ko'rinishga ega bo'lganligining eng yaxshi tirik namunalari. Ular yashaydilar koloniyalar, va ibtidoiy darajani ko'rsating uyali turli xil vazifalar uchun ixtisoslashtirish. |

Hayvonlar yoki Animalia
| Sana | Tadbir |
|---|---|
| 700–660 Ma |  Dikkinsoniya kostalari dan Ediakaran biota, 635-542 mln.yil, erta a'zosi bo'lishi mumkin Animalia. Urmetazoan: Tasvirlashi mumkin bo'lgan birinchi qoldiqlar hayvonlar ning 665 million yillik jinslarida paydo bo'ladi Trezona shakllanishi ning Janubiy Avstraliya. Ushbu qoldiqlar dastlabki gubkalar deb talqin etiladi.[7]Dan ajratish Porifera (gubkalar ) nasab.Eumetazoa /Diploblast: dan ajratish Ktenofora ("taroqli jele") nasab.Planulozoa /Paraxoxoza: dan ajratish Plakozoa va Knidariya deyarli barcha cnidarians ega asab va mushaklar. Ularga ega bo'lgan eng oddiy hayvonlar bo'lgani uchun, ularning to'g'ridan-to'g'ri ajdodlari, ehtimol asab va mushaklarni birgalikda ishlatgan birinchi hayvonlar bo'lgan. Cnidarians shuningdek, aniq tanasi va shakli aniq tanaga ega bo'lgan birinchi hayvonlardir. Ularda mavjud radial simmetriya. Birinchi ko'zlar hozirgi vaqtda rivojlangan. |
| 570–550 mln |  Proporus sp., a ksenakoelomorf. Urbilaterian:Bilateriya /Triploblastlar, Nefrozoa (555 mln.), So'nggi umumiy ajdod protostomalar (shu jumladan artropod [hasharotlar, qisqichbaqasimon] va platyzoan [yassi qurtlar] nasl-nasab) va deuterostomalar (shu jumladan umurtqali hayvonlar [odam] nasl-nasabi) .ning eng dastlabki rivojlanishi miya va of ikki tomonlama simmetriya.Bu bosqichning arxaik vakillari yassi qurtlar, uchdan hosil bo'lgan organlari bo'lgan eng oddiy hayvonlar germ qatlamlari. |
| 541 mln |  A dengiz bodringi (Aktinopyga echinitlari ), uning oziqlantiruvchi chodirlarini namoyish qilish va naycha oyoqlari. Hayvonlarning eng mashhur filasi toshqotganlik davrida dengiz turlari sifatida paydo bo'lgan Kembriya portlashi.Deuterostomalar, xordat [inson] nasabining so'nggi umumiy ajdodi, Ekinodermata (dengiz yulduzi, dengiz kirpi, dengiz bodringlari va boshqalar) va Hemichordata (Acorn qurtlari va grafolitlar ). Ushbu bosqichdan arxaik tirik qolgan bu Acorn qurti, sport a qon aylanish tizimi bilan yurak bu ham buyrak. Acorn qurtlari a gill - nafas olish uchun ishlatiladigan tuzilishga o'xshash tuzilish ibtidoiy baliq. Acorn qurtlari a pleksus ikkala dorsal va ventral asab kordlariga jamlangan. Dorsal ichak probozga etib boradi va qisman o'sha mintaqadagi epidermisdan ajratiladi. Dorsal asab shnurining bu qismi ko'pincha bo'shliq bo'lib, umurtqali hayvonlar miyasi bilan gomologik bo'lishi mumkin.[8] |
Akkordatlar
| Sana | Tadbir |
|---|---|
| 530 mln | Pikaia zamonaviy xordatlarning ikonik ajdodi va umurtqali hayvonlar.[9] Boshqa, avvalgi akkordat o'tmishdoshlari Myllokunmingiya fengjiaoa,[10] Haikouella lanceolata,[11] va Haikouichthys erkaikunensis.[12] The lancelet, hozirgi kunga qadar yashab, ibtidoiy xususiyatlarini saqlab qoladi akkordatlar. U o'xshaydi Pikaia. Konodonts erta (495 Mya va undan keyin) xordat qoldiqlarining mashhur turi; ularda katta ko'zlar, fin nurlari bilan suyaklar, chevron shaklidagi mushaklar va notoxord. Hayvon ba'zan chalkashmaslik uchun konodont, ba'zan esa konodontofor (konodont tashuvchisi) deb nomlanadi. |
| 505 mln | Birinchi umurtqali hayvonlar paydo bo'ladi: the ostrakodermalar, hozirgi zamonga oid jag'siz baliqlar lampalar va hagfishes. Haikouichthys va Myllokunmingiya bu jag'siz baliqlarning namunalari yoki Agnata. (Shuningdek qarang tarixdan oldingi baliqlar ). Ular jag'siz va ichki skeletlari xaftaga tushgan. Ularga juftlik etishmadi (ko'krak va tos suyagi) qanotlari yanada rivojlangan baliqlar. Ular avvalgilar edi Osteyxitlar (suyakli baliq).[13] |
| 480 mln | The Plakodermi edi tarixdan oldingi baliqlar. Plakodermlar jag'ning birinchi baliqlaridan biri edi (Gnathostomata ), ularning jag'lari birinchi gill kamonidan rivojlanmoqda.[14] Plasodermaning boshi va ko'krak qafasi bo'g'inli zirhli plitalar bilan yopilgan va tananing qolgan qismi masshtablangan yoki yalang'och bo'lgan. Ammo, toshbo'ron qilingan ma'lumotlar Devon tugaganidan keyin ularning avlodlarini qoldirmaganligini va akulalarga qaraganda tirik suyakli baliqlar bilan kamroq aloqadorligini ko'rsatadi.[iqtibos kerak ] |
| 410 mln |  Coelacanth 1974 yilda ushlangan |



Tetrapodlar
| Sana | Tadbir |
|---|---|
| 390 mln | Ba'zi toza suvli lobli baliqlar (Sarcopterygii ) oyoqlarini rivojlantiradi va Tetrapoda. Birinchi tetrapodlar rivojlangan sayoz va botqoqli chuchuk suv yashash joylari. Dan rivojlangan ibtidoiy tetrapodlar lobli baliq ("osteolepid" Sarcopterygian "), ikki lobli miya yassilangan bosh suyagida, keng og'izda va kalta tumshuqda, uning yuqoriga qaragan ko'zlari uning tubida yashovchi ekanligini ko'rsatib turibdi va allaqachon go'shtli poydevor bilan suyaklarning moslashuvini rivojlantirgan va suyaklar. ("Tirik qoldiq" coelacanth bog'liqdir lobli baliq Tetrapod baliqlari o'zlarining suyaklarini o'simliklar bilan bo'g'ilib qolgan sayoz suvli yashash joylarida belkurak sifatida ishlatishgan va detrit. Tirsakkacha orqaga egilgan va tizzadan oldinga egilgan orqa oyoq-qo'llarining universal tetrapod xususiyatlarini sayoz suvda yashovchi erta tetrapodlarga ishonib topshirish mumkin.[16] Panderixtis 90-130 sm (35-50 dyuym) uzunlikdagi baliqdir Devon davri (380 Mya ). U katta tetrapod o'xshash bosh. Panderixtis ko'rgazmada lobli baliqlar va erta tetrapodlar o'rtasida o'tish davri mavjud. O'xshash narsadan olingan trekvey taassurotlari Ixtiostega 's a'zolari Polshadagi dengiz oqimining cho'kindi jinslarida 390 mln. Bu tetrapod evolyutsiyasi eskirgan qoldiqlardan eski ekanligini ko'rsatadi Panderixtis orqali Ixtiostega. O'pka baliqlari erta xususiyatlarini saqlab qolish Tetrapoda. Bir misol Kvinslend o'pka baliqlari. |
| 375 mln | Tiktaalik - bu sarcopterygian (lob-finned) ko'plab tetrapodga o'xshash xususiyatlarga ega kech Devondan olingan baliqlar. Bu o'rtasidagi aniq aloqani ko'rsatadi Panderixtis va Acanthostega. |
| 365 mln | Acanthostega yo'q bo'lib ketgan amfibiya, taniqli a'zolarga ega bo'lgan birinchi hayvonlar orasida. Bu birinchilardan biri bo'lish uchun nomzod umurtqali hayvonlar quruqlikka kelish imkoniyatiga ega bo'lish. Unda bilaklar yo'q edi va umuman quruqlikda yashash uchun juda mos bo'lmagan. Oyoq-qo'llar hayvonning og'irligini ko'tarolmadi. Acanthostega ikkala o'pkasi va gillasi bor edi, shuningdek, bu lob qanotli baliqlar va quruqlikdagi umurtqali hayvonlar o'rtasidagi bog'liqlik. Ixtiostega erta tetrapod. Oyoqlari, qo'llari va barmoq suyaklari bo'lgan birinchi hayvonlardan biri bo'lib, Ixtiostega sifatida qaraladi gibrid baliq va amfibiya o'rtasida. Ixtiostega oyoqlari bo'lgan, ammo oyoq-qo'llari yurish uchun ishlatilmagan bo'lishi mumkin. Ular juda qisqa vaqtlarni suvsiz o'tkazgan bo'lsalar va oyoqlari orqali o'tib ketishgan bo'lsa kerak loy.[17] Amfibiya evolyutsiyani rivojlantirgan to'rt oyoqli birinchi hayvonlar edi Hynerpeton 360 Mya. Amfibiyalar bugungi kunda ham erta davrning ko'plab xususiyatlari saqlanib qolgan tetrapodlar. |
| 300 mln | Amfibiyalardan birinchi sudralib yuruvchilar paydo bo'ldi: Gilonomus eng qadimgi sudralib yuruvchi. Uzunligi 20 sm (8 dyuym) (dumini ham hisobga olgan holda) edi va, ehtimol, zamonaviyga o'xshaydi kaltakesaklar. Uning kichkina o'tkir tishlari bor edi va, ehtimol, egan millipedlar va erta hasharotlar. Bu kelajakning kashfiyotchisi Amniotes va sutemizuvchilarga o'xshash sudralib yuruvchilar. Alfa keratin birinchi rivojlanadi Bu yerga. U zamonaviy kaltakesaklar va qushlarning tirnoqlarida, sut emizuvchilarda esa sochlarda ishlatiladi.[18] Amniotik tuxum evolyutsiyasi Amniotani keltirib chiqaradi, sudralib yuruvchilar quruqlikda ko'payishi va quruqlikda tuxum qo'yishi mumkin. Ko'paytirish uchun ularga suvga qaytish kerak emas edi. Ushbu moslashuv ularga tog'larda birinchi marta yashash imkoniyatini berdi. Sudralib yuruvchilar, rivojlangan asab tizimiga ega amfibiyalar, o'n ikki juft bilan kranial asab. |





Sutemizuvchilar
| Sana | Tadbir |
|---|---|
| 256 mln |  Fthinosuchus, erta Terapsid Eng qadimgi sutemizuvchilarga o'xshash sudralib yuruvchilar ular pelikozavrlar. Pelikozavrlar vaqtinchalik fenestralarga ega bo'lgan birinchi hayvonlar edi. Pelikozavrlar yo'q terapevtiklar ammo tez orada ular paydo bo'ldi. Therapsida to'g'ridan-to'g'ri ajdodlari bo'lgan sutemizuvchilar. Terapsidlarda vaqtinchalik fenestralar pelikozavrlarga qaraganda kattaroq va ko'proq sutemizuvchilarga o'xshaydi, ularning tishlari ketma-ket differentsiatsiyani namoyon qiladi va keyinchalik shakllari rivojlangan ikkilamchi tanglay. Ikkilamchi tanglay hayvonga bir vaqtning o'zida ovqatlanish va nafas olishga imkon beradi va bu yanada faol, ehtimol iliq qonli hayot tarzining belgisidir.[19] |
| 220 mln | Terapsidlarning bir kichik guruhi, sinodontlar, sutemizuvchilarga o'xshash xususiyatlarni rivojlantirdi. Sinodontlarning jag'lari zamonaviy sutemizuvchilar jag'lariga o'xshaydi. Ushbu hayvonlar guruhida, ehtimol, barcha zamonaviy sutemizuvchilarning bevosita ajdodi bo'lgan tur mavjud.[20] |
| 220 mln | Kimdan Evkinodontiya (sinodontlar ) birinchi keldi sutemizuvchilar. Ko'pchilik erta sutemizuvchilar hasharotlar bilan oziqlanadigan shvedga o'xshash kichik hayvonlar edi. Qadimgi toshlarda hech qanday dalil bo'lmasa-da, ehtimol bu hayvonlarda a doimiy tana harorati va yosh bolalari uchun sut bezlari. The neokorteks miyaning mintaqasi birinchi navbatda sutemizuvchilarda rivojlangan va shu bilan ular uchun xosdir. Monotremlar zamonaviy hayvonlar orasida vakili bo'lgan sut emizuvchilarning tuxum qo'yadigan guruhidir platypus va echidna. Platypusning yaqinda genom bilan ketma-ketligi uning jinsiy genlari qushlarnikiga qaraganda qushlarga yaqinroq ekanligini ko'rsatadi Therian (jonli tug'ilish) sutemizuvchilar. Buni boshqa sutemizuvchilar bilan taqqoslaganda, birinchi bo'lib yutgan sutemizuvchilar haqida xulosa chiqarish mumkin jinsiy farqlash mavjudligi yoki etishmasligi orqali SRY geni (y-xromosomada topilgan) monotreme nasab bo'linib ketganidan keyin rivojlandi. |
| 160 mln | Juramaia sinensis[21] eng qadimgi evteriya sutemizuvchilarning qoldiqlari. |
| 100 mln | Oxirgi umumiy ajdod ning sichqonlar va odamlar (qoplamaning asosi) Evarxontoglires ). |



Primatlar
| Sana | Tadbir |
|---|---|
| 85-66 mln | Kichkina, tungi, daraxtzor, hasharotlarni iste'mol qiladigan sutemizuvchilar guruhi chaqirildi Euarchonta ga olib keladigan spetsifikatsiyani boshlaydi buyurtmalar ning primatlar, xiyonat qiluvchilar va uchadigan lemurlar. Primatomorfa Evarxontaning bo'linmasi bo'lib, u primatlar va ularning ajdodlari kelib chiqqan primatlarni o'z ichiga oladi Plesiadapiformes. Dastlabki novda, Plesiadapis, hali ham bosh tomonida tirnoqlari va ko'zlari bor edi, bu daraxtlarga qaraganda tezroq erga tushar edi, lekin u uzoq vaqt davomida pastki shoxlarga, mevalar va barglar bilan oziqlana boshladi. Plesiadapiformes, ehtimol barcha primatlarning ajdod turlarini o'z ichiga oladi.[22] Ular birinchi bo'lib fotoalbomlarda 66 million yil oldin paydo bo'lgan Bo'r-paleogen yo'q bo'lib ketish hodisasi bu dinozavrlarning aksariyatini, shu jumladan Yerdagi o'simlik va hayvonot turlarining to'rtdan uch qismini yo'q qildi.[23][24] Oxirgi Plesiadapiformes biri Karpolestlar simpsoni, raqamlarni tushunadigan, ammo ko'zlari oldinga emas. |
| 63 mln | Primatlar subordinatsiyalarga bo'linish Strepsirrhini (ho'l burunli primatlar) va Xaplorrin (quruq burunli primatlar). Strepsirrin tarkibida ko'pi bor prokurorlar; zamonaviy misollar kiradi lemurlar va lorises. Gaplorrinlarga ikkita tirik guruh kiradi: prosimian buzadigan amallar va simian maymunlar, shu jumladan maymunlar. Eng qadimgi haplorrinlardan biri Teilhardina asiatica, kichkina ko'zlari bilan sichqoncha kattaligi, kunduzgi mavjudot. Haplorrhini metabolizmi ishlab chiqarish qobiliyatini yo'qotdi S vitamini, barcha avlodlarni S vitamini o'z ichiga olgan mevalarni ratsioniga kiritishga majbur qilish. |
| 30 mln | Xaplorrin buzilishlarga bo'linadi Platirrini va Katarrin. Platirrinlar, Yangi dunyo maymunlari, ularning quyruqlari oldindan tarqalgan, erkaklar esa ko'r-ko'rona. Avlodlari Platirriniga aylanadigan shaxslar Janubiy Amerikaga yoki a o‘simliklar sal yoki a orqali quruqlik ko'prigi (gipoteza hozir ma'qul[25]). Katarrinlar asosan ichida qolishgan Afrika ikki qit'a bir-biridan uzoqlashganda. Katarrinlarning mumkin bo'lgan dastlabki ajdodlari kiradi Misropitek va Saadanius. |
| 25 mln | Katarrin ikkita superfililaga bo'linadi, Qadimgi dunyo maymunlari (Cercopithecoidea) va maymunlar (Gominoideya ). Bizning trikromatik rang ko'rish bu davrda genetik kelib chiqishiga ega edi. Prokonsul katarinli primatlarning dastlabki turi edi. Ularning aralashmasi bor edi Qadimgi dunyo maymuni va maymun xususiyatlari. Prokonsul 's maymun o'xshash xususiyatlarga ingichka kiradi tish emal, tor ko'krak va kalta oyoq oyoqlari bilan engil qurilish va daraxt to'rtburchak hayot tarzi. Uning maymunga o'xshash xususiyatlari - bu dumining etishmasligi, maymunga o'xshash tirsaklar va tana hajmiga nisbatan biroz kattaroq miya. Proconsul africanus maymunlarning ham, mayda maymunlarning ham, shu jumladan odamlarning ajdodidir. |




Hominidae
| Sana | Tadbir |
|---|---|
| 18 mln | Hominidae (buyuk maymun ajdodlari) ning ajdodlaridan aniqlang gibbon (kichik maymunlar) v. 20 dan 16 gacha.[26] |
| 16 mln | Gomininlar ajdodlar ajdodlaridan ajrata olishadi orangutan v orasida. 18 dan 14 gacha.[27] Pierolapithecus catalaunicus a deb o'ylashadi umumiy ajdod odamlar va boshqa buyuk maymunlar, yoki hech bo'lmaganda avvalgi qazilma topilmalaridan ko'ra bizni umumiy ajdodga yaqinlashtiradigan tur. U hozirgi odamlar va boshqa buyuk maymunlar singari daraxtlarga chiqish uchun maxsus moslashuvlarga ega edi: keng, tekis ko'krak qafasi, qattiq pastroq umurtqa pog'onasi, egiluvchan bilaklar va elka pichoqlari uning orqa tomonida joylashgan. |
| 12 mln | Danuvius guggenmosi saqlanib qolgan, birinchi kashf etilgan kech miosen maymunidir uzun suyaklar va zamonaviy maymunlarning anatomik tuzilishi va harakatlanishini juda yaxshi yoritib beradi.[28] Ikkala daraxtga osilganligi uchun moslashtirildi (shubhali xatti-harakatlar ) va ikki oyoq bilan yurish (bipedalizm ) - aksincha, hozirgi gominidlar orasida odamlar ikkinchisiga, boshqalari esa birinchisiga yaxshi moslashgan. Danuvius ilgari ma'lum bo'lgan maymundan farqli o'laroq harakatlanish usuli "kengaytirilgan oyoq-qo'llarni qistirmoq" deb nomlangan, to'g'ridan-to'g'ri daraxt shoxlari bo'ylab yurgan va o'zini to'xtatib turish uchun qurol ishlatgan. Odamlar va boshqa maymunlar orasidagi so'nggi umumiy ajdod, xuddi shunday harakatlanish uslubiga ega bo'lgan. |
| 10 mln | Hozirgi vaqtda odamlar va nasl tomonidan namoyish etilayotgan qoplama Pan (oddiy shimpanzeler va bonobos ) ajdodlaridan ajratilgan gorilla v orasida. 10 dan 8 gacha.[29] |
| 6 mln | Gominini: The so'nggi umumiy ajdod ning odamlar va shimpanze taxminan 10 dan 5 million yilgacha yashagan deb taxmin qilinadi. Ham shimpanzalarda, ham odamlarda a gırtlak hayotning dastlabki ikki yilidagi repozitsiyalar tomoq va o'pka, bu umumiy ajdodlarimizning bu xususiyatga ega ekanligini ko'rsatib, odamlarda ovozli nutq uchun zarur shartdir. Belgilanish 10 Madan ko'p o'tmay boshlangan bo'lishi mumkin, ammo nasablar orasidagi kechki aralashma 5 Ma dan keyin sodir bo'lishi mumkin. Nomzodlari Gominina yoki Gomininlar shu davrda yashagan turlarga kiradiOuranopitekus (taxminan 8 mln.),Greekopitek (taxminan 7 mln.),Sahelanthropus tchadensis (taxminan 7 mln.),Orrorin tugenensis (taxminan 6 mln.). Ardipitek juda erta yoki bo'lishi mumkin hominin tur (qabila Gominini va obuna bo'lish Gominina ). Adabiyotda ikkita tur tasvirlangan: A. ramidus, taxminan 4,4 million yil oldin yashagan[30] erta davrida Plyotsen va A. kadabba, taxminan 5,6 million yil oldin tuzilgan[31] (kech Miosen ). A. ramidus 300 dan 350 sm gacha bo'lgan kichik miyaga ega edi3. Bu zamonaviy bilan bir xil darajada bonobo va ayol oddiy shimpanze miya; u shunga o'xshash avstralopitektsiyalarning miyasidan biroz kichikroq Lyusi (400 dan 550 sm gacha)3) va zamonaviyning beshdan bir qismidan biroz kattaroq Homo sapiens miya.Ardipithecus daraxt edi, ya'ni u asosan o'rmonda yashagan, u erda boshqa o'rmon hayvonlari bilan raqobatlashadigan, shubhasiz, shimpanzalarning zamonaviy ajdodi ham bo'lgan. Ardipitek, ehtimol ikki oyoqli uning kosasi shaklidagi tos suyagi, uning burchagi foramen magnum Bilagi suyaklari ham ingichka, ammo oyoqlari hali uzoq masofalarga yurishdan ko'ra ushlashga moslashgan edi. |
| 3.6 mln | A'zosi Australopithecus afarensis vulkanik kulda odamga o'xshash oyoq izlarini qoldirdi Laetoli, shimoliy Tanzaniya, to'la vaqtli bipedalizmning kuchli dalillarini taqdim etish. Australopithecus afarensis 3.9 va 2.9 million yil oldin yashagan va eng qadimgi davrlardan biri hisoblanadi homininlar - rivojlangan va nasabni o'z ichiga olgan ushbu turlar Homo va Homo'shimpanzelar qatoridan ajralib chiqqanidan keyin eng yaqin qarindoshlari. Bu shunday deb o'ylashadi A. afarensis ikkala naslga ham ajdodlardan bo'lgan Avstralopitek va tur Homo. Zamonaviy va yo'q bo'lib ketgan buyuk bilan taqqoslaganda maymunlar, A. afarensis tish va tishlar kamaygan edi, garchi ular zamonaviy odamlarga qaraganda ancha kattaroq edi. A. afarensis shuningdek, nisbatan kichik miya kattaligi (380-430 sm and) va prognatik (oldingi proektsion) yuzga ega. Savolannya muhitida avstralopitektsiyalar topilgan; ular, ehtimol, parhezni maydalangan go'shtni kiritish uchun ishlab chiqdilar. Ning tahlillari Australopithecus africanus pastki umurtqalar homiladorlik paytida ham bipedalizmni qo'llab-quvvatlash uchun bu suyaklar ayollarda o'zgarganligini ko'rsatadi. |
| 3,5-3,3 mln | Kenyanthropus platyops, mumkin bo'lgan ajdodi Homo, dan chiqadi Avstralopitek. Tosh qurollari ataylab qurilgan.[32] |
| 3 mln | Ikki oyoqli avstralopitekinlar (subtribe bir turi) Gominina ) savannalarida rivojlanib boradi Afrika tomonidan ov qilinmoqda Megantereon. Yo'qotish tana sochlari to'la rivojlanishiga parallel ravishda 3 dan 2 mln. gacha sodir bo'ladi bipedalizm.[33] |

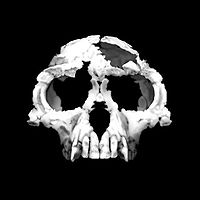
Homo
| Sana | Tadbir |
|---|---|
| 2,5-2,0 mln | Erta Homo dan iborat bo'lgan Sharqiy Afrikada paydo bo'ladi avstralopiteksin ajdodlar tosh qurollar ning boshini belgilang Quyi paleolit.Australopithecus garhi taxminan 2,5 mln. tosh toshlardan foydalangan.Homo habilis nomi berilgan eng qadimgi turlardir Homo, tomonidan Liki va boshq. (1964).H. habilis orasidagi oraliqdir Australopithecus afarensis va H. erectus va uni turkum ichida qayta tasniflash bo'yicha takliflar mavjud Avstralopitek, kabi Australopithecus habilis. Da topilgan tosh qurollar Shanxen Xitoyda joylashgan va 2,12 million yil ilgari qurilgan sayt Afrikadan tashqarida gomininlarning eng qadimgi dalillari hisoblanib, Gruziyadagi Dmanisidan 300 ming yilga oshib ketgan.[34] |
| 1,9-0,5 mln | Homo erectus erta kelib chiqadi Homo yoki kech Avstralopitek. Homo habilis, anatomiya va fiziologiyadan sezilarli farq qilsa-da, ajdodi deb o'ylashadi Homo ergaster yoki afrikalik Homo erectus; lekin u bilan birga yashaganligi ham ma'lum H. erectus deyarli yarim million yil davomida (taxminan 1,5 mln. gacha). Uning paydo bo'lishidan taxminan 1,9 mln. H. erectus Sharqiy Afrika va Janubi-G'arbiy Osiyoda tarqalgan (Homo georgicus ). H. erectus rivojlangan ma'lum bo'lgan birinchi tur olovni boshqarish, taxminan 1,5 mln. H. erectus keyinchalik ko'chib ketadi Evroosiyo bo'ylab, Janubi-Sharqiy Osiyo ga 0,7 mln. ga etib boradi, bu bir qatorda tasvirlangan pastki turlari.[35] Qorong'u terining rivojlanishi taxminan 1,2 mln.[36] Homo salafi odamlar va neandertallarning umumiy ajdodi bo'lishi mumkin.[37][38] Hozirgi hisob-kitoblarga ko'ra, odamlarda taxminan 20000-25000 kishi bor genlar va ularning 99% ulushiga ega DNK endi yo'q bo'lib ketgan Neandertal [39] va ularning 95-99% DNK ularning eng yaqin yashaydigan evolyutsion qarindoshi bilan shimpanze.[40][41] Ning insoniy varianti FOXP2 gen (nutqni boshqarish bilan bog'liq) neandertallarda bir xil ekanligi aniqlandi.[42] |
| 0,8-0,3 mln | Tafovut Neandertal va Denisovan umumiy ajdodlardan kelib chiqqan nasablar.[43]Homo heidelbergensis (Afrikada ham nomi bilan tanilgan Homo rodesiensis ) uzoq vaqtdan beri neandertal va zamonaviy inson nasablarining so'nggi umumiy ajdodiga nomzod bo'lishi mumkin deb o'ylagan edi, ammo genetik dalillar Sima de los Huesos 2016 yilda nashr etilgan fotoalbomlar shuni ko'rsatadiki H. heidelbergensis butunlay neandertal nasabiga "neandertalgacha" yoki "erta neandertal" sifatida kiritilishi kerak, neandertal va zamonaviy nasllar o'rtasidagi ixtilof vaqti paydo bo'lishidan oldin orqaga surilgan. H. heidelbergensis, taxminan 600000 dan 800000 yil oldin, taxminan yoshi Homo salafi.[44][45] Qattiq oyoq izlari taxminan 350 ka ga tegishli va u bilan bog'liq H. heidelbergensis 2003 yilda Italiyaning janubida topilgan.[46] |
Homo sapiens
| Sana | Tadbir |
|---|---|
| 300-130 ka |  Qayta qurish H. neandertalensis Qoldiqlar tegishli H. sapiens, da topilgan tosh qurollar bilan bir qatorda, taxminan 300000 yil ilgari topilgan Jebel Irhoud, Marokash[47] uchun eng qadimiy qazilma dalillarni berish anatomik jihatdan zamonaviy Homo sapiens Insonlarning zamonaviy ishtiroki Sharqiy Afrika (Gademotta ), 276 kya da.[48]2017 yilda Isroilda topilgan 177 ming yillik jag 'suyagi qoldig'i Afrika tashqarisida topilgan eng qadimgi odam qoldiqlari hisoblanadi.[49] Biroq, 2019 yil iyul oyida antropologlar 210 ming yillik qoldiqlari topilganligi haqida xabar berishdi H. sapiens va 170 ming yillik qoldiqlari H. neandertalensis yilda Apidima g'ori, Peloponnes, Gretsiya, avvalgisidan 150 ming yoshdan katta H. sapiens Evropada topilgan narsalar.[50][51][52] Neandertallar dan chiqmoq Homo heidelbergensis bir vaqtning o'zida nasab (300 ka). Patrilineal va matrilineal tirik odamlarning so'nggi umumiy ajdodlari (MRCA) taxminan 200 dan 100 ka gacha[53][54]Patrilineal MRCA bo'yicha ba'zi taxminlar bilan bir oz yuqoriroq, 250 dan 500 kya gacha.[55] 160,000 yil oldin, Homo sapiens idaltu ichida Avash daryosi vodiysi (hozirgi zamonga yaqin) Herto qishlog'i, Efiopiya) mashq qilgan mujassamlash.[56] |
| 130-80 ka | Dengiz izotopi 5-bosqich (Eemian ). Zamonaviy insonning mavjudligi Janubiy Afrika va G'arbiy Afrika.[57]Mitokondriyal haplogroupning ko'rinishi (mt-haplogroup) L2. |
| 80-50 ka | MIS 4, ning boshi Yuqori paleolit. Uchun dastlabki dalillar xatti-harakatlarning zamonaviyligi.[58]MT-haplogrouplarning ko'rinishi M va N. Janubiy tarqalish Afrikadan tashqariga ko'chish, Proto-avstraloid Okeaniya orollari.[59] Arxaik aralashma dan Neandertallar Evrosiyoda,[60][61] dan Denisovaliklar Sharqiy Evrosiyoda oz miqdordagi Okeaniyada,[62] va Sahroi Afrikadagi arxaik odamlarning aniqlanmagan afrikalik nasabidan, shuningdek Osiyo va Okeaniyada neandertal va denisovanlarning nasldan naslga oid turlari.[63][64][65][66] |
| 50-25 ka |  Qayta qurish Oase 2 (taxminan 40 ka) Xulq-atvor zamonaviyligi ga ko'ra rivojlanadi "oldinga katta sakrash" nazariya.[67]Yo'qolib ketish Homo floresiensis.[68]M168 mutatsiya (barcha afrikalik bo'lmagan erkaklar tomonidan olib boriladi) .mt-haplogruplarning ko'rinishi U va K.Evropada Peopling, Shimoliy Osiyo aholisi Mamont dasht. Paleolit san'ati.Neandertallarning yo'q bo'lib ketishi va insonning boshqa arxaik variantlari (mumkin bo'lgan omon qolish bilan) gibrid Osiyo va Afrikadagi populyatsiyalar.) Y-Haplogroupning ko'rinishi R2; mt-haplogrouplar J va X. |
| 25 ka dan keyin | Oxirgi muzlik maksimal darajasi; Epipaleolit / Mezolit / Golotsen.Amerika qit'asi poytaxtlari.Tashqi ko'rinishi: Y-Haplogroup R1a; mt-haplogrouplar V va T.Har xil yaqinda yuzaga kelgan kelishmovchilik atrof-muhit bosimi bilan bog'liq, masalan. engil teri ichkarida Evropaliklar va Sharqiy osiyoliklar (KITLG, ASIP ), 30 ka dan keyin;[69]Inuit yuqori yog'li dietaga va sovuq iqlimga moslashish, 20 ka.[70] Kech tirik qolganlarning yo'q bo'lib ketishi arxaik odamlar boshida Golotsen (12 ka) .da qatnashadigan populyatsiyalardagi selektsiya bosimi tufayli tezlashib ketgan farq Neolitik inqilob 12 ka dan keyin, masalan, Sharqiy Osiyo turlari ADH1B bilan bog'liq guruchni xonakilashtirish,[71] yoki laktaza doimiyligi.[72][73] |
Shuningdek qarang
Adabiyotlar
- ^ Finarelli, J.A .; Klayd, Vashington (2004). "Gominoid filogeniyani qayta baholash: morfologik va vaqtinchalik ma'lumotlarning muvofiqligini baholash". Paleobiologiya. 30 (4): 614.
- ^ Patterson N, Rixter DJ, Gnerre S, Lander ES, Reyx D (2006). "Odamlar va chimpanzaklarning kompleks spetsifikatsiyasi uchun genetik dalillar". Tabiat. 441 (7097): 1103–08. Bibcode:2006 yil natur.441.1103P. doi:10.1038 / nature04789. PMID 16710306. S2CID 2325560.
- ^ ning tasnifiga qarab Homo heidelbergensis nasab; 0.8, agar neandertallar sifatida tasniflangan bo'lsa H. sapiens neanderthalensisyoki agar bo'lsa H. sapiens dan divergentsiyadan kladistik ravishda aniqlanadi H. neandertalensis, Mavjud qazilma dalillarga asoslanib 0,3.
- ^ "" Jinsiy aloqada bo'lgan tajribalarni o'tkazish juda qiyin bo'lgan, - dedi Goddard. - Eksperimentda odam qiziqish jihatini hisobga olmaganda, hamma narsani doimiy ravishda ushlab turishi kerak. Demak, hech qanday yuqori organizmlardan foydalanish mumkin emas, chunki ular kerak ko'payish uchun jinsiy aloqada bo'ling va shuning uchun aseksual nazoratni ta'minlamang. '
Goddard va uning hamkasblari jinsiy aloqa populyatsiyalarning yangi sharoitlarga tezroq moslashishiga jinssiz populyatsiyalarga imkon beradi degan g'oyani sinab ko'rish uchun bir hujayrali organizmga xamirturushga murojaat qilishdi." Jinsiy aloqa evolyutsiyani tezlashtiradi, o'rganish natijalari (URL manziliga 2005 yil 9 yanvarda kirilgan) - ^ Dawkins, R. (2005), Ajdodlar haqidagi ertak: Evolyutsiya tongiga ziyorat, Xyuton Mifflin Harkort, ISBN 978-0-618-61916-0
- ^ "Proterospongia - bu nodir chuchuk suv protisti, Choanoflagellata kolonial a'zosi." "Proterospongia o'zi gubkalarning ajdodi emas. Ammo u gubkalar va boshqa metazoanlar ajdodi qanday bo'lishi mumkinligi uchun foydali namuna bo'lib xizmat qiladi." http://www.ucmp.berkeley.edu/protista/proterospongia.html Berkli universiteti
- ^ Maluf, Adam S.; Rose, Ketrin V.; Plyaj, Robert; Samuels, Bredli M.; Kalmet, Kler S.; Ervin, Duglas X.; Pueri, Jerald R.; Yao, Nan; Simons, Frederik J. (2010 yil 17-avgust). "Janubiy Avstraliyadan Marinoga qadar bo'lgan ohaktoshlarda mumkin bo'lgan hayvon tanasi qoldiqlari". Tabiatshunoslik. 3 (9): 653–59. Bibcode:2010 yil NatGe ... 3..653M. doi:10.1038 / ngeo934. S2CID 13171894.
- ^ Barns, Robert D. (1982). Umurtqasizlar zoologiyasi. Filadelfiya: Xolt-Sonders xalqaro. 1018-26 betlar. ISBN 978-0-03-056747-6.
- ^ "Shubhasiz, umurtqali hayvonlarning ajdodlari Kembriyada yashagan bo'lishi kerak, ammo ular haqiqiy umurtqali hayvonlar - protoxordatlarning umurtqasiz kashshoflari deb taxmin qilingan. Pikaia eng qadimiy fotoalbom protokordati sifatida katta targ'ib qilingan. " Richard Dokkins 2004 Ajdodlar ertagi p. 289, ISBN 0-618-00583-8
- ^ Shu, D.G .; Luo, H.L .; Konvey Morris, S .; Chjan, X. L .; Xu, S.X .; Chen, L .; Xan, J .; Chju, M .; Li, Y .; Chen, L.Z. (1999). "Janubiy Xitoydan pastki Kembriya umurtqali hayvonlari". Tabiat. 402 (6757): 42–46. Bibcode:1999 yil Nat.402 ... 42S. doi:10.1038/46965. S2CID 4402854.
- ^ Chen, J.Y .; Xuang, D.Y .; Li, CW (1999). "Dastlabki kembriy kraniatiga o'xshash xordat". Tabiat. 402 (6761): 518–22. Bibcode:1999 yil Natur.402..518C. doi:10.1038/990080. S2CID 24895681.
- ^ Shu, D.-G.; Morris, S.Konvey; Xan, J .; Chjan, Z.-F .; Yasui, K .; Yanvier, P .; Chen, L .; Chjan, X.-L .; Liu, J.-N .; Li, Y .; Liu, H.-Q. (2003 yil yanvar). "Erta Kembriy umurtqali haykuytisning boshi va umurtqasi". Tabiat. 421 (6922): 526–529. Bibcode:2003 yil Noyabr.421..526S. doi:10.1038 / nature01264. PMID 12556891. S2CID 4401274.
- ^ Ushbu birinchi umurtqali hayvonlar tirik xagfish va chiriyotgan kabi jag'larga ega emas edilar. Jawed umurtqali hayvonlar 100 million yildan so'ng, Siluriyada paydo bo'lgan. http://www.ucmp.berkeley.edu/vertebrates/vertintro.html Berkli universiteti
- ^ "Birinchi gill kamarining suyaklari yuqori va pastki jag'larga aylandi." (Rasm)
- ^ Ma'lumotlar qatlami 410-da topilgan toshbaqa jilosi mya Buxan yaqinida to'plangan Viktoriya, Avstraliya Sharqiy Gippslend, hozirgi paytda eng keksa seelant bo'yicha rekordchiga ega; unga ism berilgan Eoactinistia foreyi 2006 yil sentyabr oyida nashr etilganida. [1]
- ^ "O'pka baliqlari tetrapodlarning eng yaqin yashaydigan qarindoshlari va ular bilan bir qator muhim xususiyatlarni baham ko'rishadi. Bu belgilar orasida tish emallari, o'pka qon oqimini tana qon oqimidan ajratish, bosh suyagi suyaklarini joylashishi va mavjudligi to'rtta tetrapod oyoqlari bilan bir xil holatga va tuzilishga ega bo'lgan to'rtta o'xshash o'lchamdagi a'zolar. " http://www.ucmp.berkeley.edu/vertebrates/sarco/dipnoi.html Berkli universiteti
- ^ "amfibiyalar sudralib yuruvchilar va o'zimiz bilan baham ko'radigan ajdodimizmi?" "Bu ehtimol o'tish davri osti qoldiqlari, ular orasida juda ko'p o'rganilgan Acanthostega, bu butunlay suvda bo'lgan ko'rinadi va Ixtiostega" Richard Dokkins 2004 Ajdodlar ertagi p. 250, ISBN 0-618-00583-8
- ^ Ekxart, L .; Valle, L. D .; Jeyger, K .; Balaun, C .; Sabo, S .; Nardi, A .; Byuxberger M.; Hermann, M .; Alibardi, L .; Tschachler, E. (2008 yil 10-noyabr). "Soch keratiniga o'xshash oqsillarni kodlovchi sudralib yuruvchilar genlarini aniqlash sochlarning evolyutsion kelib chiqishining yangi stsenariysini taklif qiladi". Milliy fanlar akademiyasi materiallari. 105 (47): 18419–18423. doi:10.1073 / pnas.0805154105. PMC 2587626. PMID 19001262.
- ^ "Ko'p jihatdan pelikozavrlar sudralib yuruvchilar va sutemizuvchilar o'rtasida oraliqdir" http://www.ucmp.berkeley.edu/synapsids/pelycosaurs.html Berkli universiteti
- ^ "Thrinaxodon, har qanday qoldiq kabi, ajdodning o'zi emas, balki ajdodimizning amakivachchasi deb o'ylash kerak. U sutemizuvchilarga o'xshash sudralib yuruvchilar guruhining a'zosi edi. Sinodontlar shunchalik sutemizuvchilarga o'xshar edilar, bu jozibali. "ularni sutemizuvchilar deb atash. Ammo biz ularni nima deyishimiz kimga ahamiyat beradi? Ular deyarli mukammal qidiruv vositalar". Richard Dokkins 2004 Ajdodlar ertagi p. 211, ISBN 0-618-00583-8
- ^ Luo, ZX; Yuan, CX; Men, QJ; Ji, Q (avgust 2011). "Yuraning evteriya sutemizuvchisi va marsupials va platsentallarning divergensiyasi". Tabiat. 476 (7361): 442–45. Bibcode:2011 yil natur.476..442L. doi:10.1038 / tabiat10291. PMID 21866158. S2CID 205225806.
- ^ "Concestor 8 qanday bo'lganini qayta tiklashga yordam beradigan toshqotganliklarga plesiadapi shakllari deb nomlangan katta guruh kiradi. Ular o'z vaqtida yashashgan va ular barcha primatlarning buyuk ajdodlaridan kutgan fazilatlarga ega" Richard Dokkins 2004 Ajdodlar ertagi p. 136, ISBN 0-618-00583-8
- ^ Renne, Pol R.; Deino, Alan L.; Xilgen, Frederik J.; Kayper, Klaudiya F.; Mark, Darren F.; Mitchell, Uilyam S.; Morgan, Liya E.; Mundil, Roland; Smit, yanvar (2013 yil 7-fevral). "Bo'r-paleogen chegarasi atrofidagi muhim voqealarning vaqt o'lchovlari". Ilm-fan. 339 (6120): 684–87. Bibcode:2013 yil ... 339..684R. doi:10.1126 / fan.1230492. PMID 23393261. S2CID 6112274.
- ^ "Paleontologlar eng ibtidoiy primat skeletini topdilar", Phys.org (2007 yil 23-yanvar).
- ^ Alan de Keyrush, Maymunning sayohati, Asosiy kitoblar, 2014 y.
- ^ "Gominoidlar daraxtining ildizida yangi primat turi". phys.org. Olingan 2020-05-29.
- ^ Raauma, Rayan; Sternera, K (2005). "To'liq mitoxondriyal genomlardan taxmin qilingan katarrin primat divergentsiyasi sanalari" (PDF). Inson evolyutsiyasi jurnali. 48 (3): 237–57. doi:10.1016 / j.jhevol.2004.11.007. PMID 15737392.
- ^ Bohme, Madelayn; Spassov, Nikolay; Fuss, Xoxen; Trosher, Adrian; Din, Endryu S.; Prieto, Jerom; Kirsher, Uve; Lechner, Tomas; Boshlandi, Devid R. (noyabr 2019). "Buyuk maymunlar va odamlarning ajdodidagi yangi miosen maymuni va harakatlanish". Tabiat. 575 (7783): 489–493. Bibcode:2019 yil natur.575..489B. doi:10.1038 / s41586-019-1731-0. ISSN 1476-4687. PMID 31695194. S2CID 207888156.
- ^ Popadin, Konstantin; Gunbin, Konstantin; Peshkin, Leonid; Annis, Sofiya; Fleyshman, Zoe; Kraytsberg, Genya; Markuzon, Natalya; Akkermann, Rebekka R.; Xrapko, Konstantin (2017-10-19). "Mitoxondriyal psevdogenlar gominid evolyutsiyasida turlararo takroriy duragaylanishni taklif qiladi". bioRxiv: 134502. doi:10.1101/134502.
- ^ Perlman, Devid (2001 yil 12-iyul). "Efiopiyadan topilgan qoldiqlar insonning eng qadimgi ajdodi bo'lishi mumkin". National Geographic News.
Boshqa hammualliflardan biri - D. D. Uayt, UC-Berkeleyning paleoantropologi, u 1994 yilda insoniyatgacha Ardipithecus ramidus nomli 4,4 million yil ichida eng qadimgi tosh qoldiqlarini topdi.
- ^ Oq, Tim D.; Asfav, Berxan; Beyene, Yonas; Xayl-Selassi, Yoxannes; Lovejoy, C. Ouen; Suva, Gen; WoldeGabriel, Giday (2009). "Ardipithecus ramidus va erta gominidlarning paleobiologiyasi ". Ilm-fan. 326 (5949): 75–86. Bibcode:2009 yil ... 326 ... 75 Vt. doi:10.1126 / science.1175802. PMID 19810190. S2CID 20189444.
- ^ Xarmand, Soniya; Lyuis, Jeyson E.; Feybel, Kreyg S.; Lepre, Kristofer J.; Prat, Sandrin; Lenobl, Arno; Boes, Xavier; Kvinn, Rhonda L.; Brenet, Mishel; Arroyo, Adrian; Teylor, Nikolay; Klement, Sofi; Daver, Giyom; Brugal, Jan-Filipp; Liki, Luiza; Mortlok, Richard A.; Rayt, Jeyms D .; Lokorodi, Sammy; Kirva, Kristofer; Kent, Dennis V.; Roche, Helen (2015). "Lomekwi 3, G'arbiy Turkana, Keniyadan 3,3 million yillik tosh qurollar". Tabiat. 521 (7552): 310–15. Bibcode:2015 yil Noyabr 521..310H. doi:10.1038 / tabiat14464. PMID 25993961. S2CID 1207285.
- ^ Rukston, Grem D.; Uilkinson, Devid M. (2011-12-27). "Gomininlarda sochlarning to'kilishi va ikki oyoqliligi uchun qizib ketish va tanlovni oldini olish". Milliy fanlar akademiyasi materiallari. 108 (52): 20965–20969. Bibcode:2011PNAS..10820965R. doi:10.1073 / pnas.1113915108. ISSN 0027-8424. PMC 3248486. PMID 22160694.
- ^ Chju, Chjaoy; Dennell, Robin; Xuang, Veyven; Vu, Yi; Tsyu, Shifan; Yang, Shixia; Rao, Jiguo; Xou, Yamey; Xie, Dzyubing; Xan, Tszayvey; Ouyang, Tingping (2018). "Taxminan 2,1 million yil ilgari Xitoyning Loess platosidagi Gominin okkupatsiyasi". Tabiat. 559 (7715): 608–12. Bibcode:2018Natur.559..608Z. doi:10.1038 / s41586-018-0299-4. PMID 29995848. S2CID 49670311.
- ^ NOVA: Inson bo'lish 2-qism http://video.pbs.org/video/1319997127/
- ^ Jablonski, Nina G. (2004 yil oktyabr). "Inson terisi va teri rangining evolyutsiyasi". Antropologiyaning yillik sharhi. 33 (1): 585–623. doi:10.1146 / annurev.anthro.33.070203.143955. S2CID 53481281.
- ^ Bermudez de Kastro, J. M. (1997 yil 30-may). "Atapuerkaning pastki pleystotsenidan Ispaniya, Ispaniya: neandertallar va zamonaviy odamlarga ajdod". Ilm-fan. 276 (5317): 1392–1395. doi:10.1126 / science.276.5317.1392. PMID 9162001.
- ^ Yashil, Richard E.; Krause, Yoxannes; Ptak, Syuzan E .; Briggs, Adrian V.; Ronan, Maykl T.; Simons, Yan F.; Du, Ley; Egxolm, Maykl; Rotberg, Jonathan M.; Paunovich, Maja; Pääbo, Svante (2006 yil noyabr). "Neandertal DNKning bir million tayanch juftligini tahlil qilish". Tabiat. 444 (7117): 330–336. Bibcode:2006 yil natur.444..330G. doi:10.1038 / nature05336. PMID 17108958. S2CID 4320907.
- ^ "Rubinning aytishicha, hozirgacha o'tkazilgan tahlillarga ko'ra inson va neandertal DNKlari 99,5 foizdan 99,9 foizgacha bir xil." Neandertal suyagi DNK haqida ma'lumot beradi (URL manzilga 2006 yil 16-noyabrda kirilgan)
- ^ "Xulosa shuni anglatadiki, biz DNK ketma-ketligining 98,5 foizini shimpanze bilan bo'lishamiz, ehtimol bu xato. Ushbu namuna uchun 95% asosiy juftlarning shimpanze va inson DNKlari o'rtasida to'liq bo'lishini taxmin qilish mumkin." Britten, R.J. (2002). "Shimpanze namunalari va odamning DNK sekanslari o'rtasidagi farq 5% ni tashkil etadi, indellarni hisobga olganda". PNAS. 99 (21): 13633–35. Bibcode:2002 yil PNAS ... 9913633B. doi:10.1073 / pnas.172510699. PMC 129726. PMID 12368483.
- ^ "... inson genomini tashkil etuvchi uch milliard harfdan atigi 15 millioni - 1 foizga ham etmaydi - olti million yil ichida yoki odam va chimildiq nasllari bir-biridan ajralib turgandan beri o'zgargan." Pollard, K.S. (2009). "Bizni nima odam qiladi?". Ilmiy Amerika. 300–5 (5): 44–49. Bibcode:2009SciAm.300e..44P. doi:10.1038 / Scientificamerican0509-44. PMID 19438048. S2CID 38866839.
- ^ Krauz J; Lalueza-Fox C; Orlando L; Enard V; Yashil RE; Burbano XA; Xublin JJ; Hanni C; Fortea J; de la Rasilla M; Bertranpetit J; Rosas A; Pääbo S (2007 yil noyabr). "Zamonaviy odamlarning olingan FOXP2 varianti Neandertals bilan bo'lishdi". Curr. Biol. 17 (21): 1908–12. doi:10.1016 / j.cub.2007.10.008. hdl:11858 / 00-001M-0000-000F-FED3-1. PMID 17949978. S2CID 9518208. Xulosa – Nyu-York Tayms (2007-10-19).
- ^ Stein, Richard A. (oktyabr 2015). "Raqamni tahlil qilish qo'shish uchun boshlanadi". Genetik muhandislik va biotexnologiya yangiliklari. 35 (17): 20, 22–23. doi:10.1089 / gen.35.17.09.
- ^ Meyer, Matias; Arsuaga, Xuan-Luis; De Filippo, Sezar; Nagel, Sara; Aximu-Petri, Ayinuer; Nikel, Birgit; Martines, Ignasio; Grasiya, Ana; De Kastro, Xose Mariya Bermudes; Karbonell, Eudald; Viola, Bens; Kelso, Janet; Prüfer, Kay; Pääbo, Svante (mart 2016). "O'rta pleystotsen Sima de los Huesos homininlaridan yadroli DNK sekanslari". Tabiat. 531 (7595): 504–07. Bibcode:2016Natur.531..504M. doi:10.1038 / tabiat17405. PMID 26976447. S2CID 4467094.
- ^ Callaway, Ewen (2016). "Neandertallar paydo bo'lgan qadimgi odamlarning eng qadimgi DNK tafsilotlari". Tabiat. 531 (7594): 296–86. Bibcode:2016 yil natur.531..296C. doi:10.1038 / 531286a. PMID 26983523. S2CID 4459329.
- ^ Mietto, Paolo; Avanzini, Marko; Rolandi, Juzeppe (2003). "Paleontologiya: Pleystotsen vulkanik kulida odam izlari". Tabiat. 422 (6928): 133. Bibcode:2003 yil natur.422..133M. doi:10.1038 / 422133a. PMID 12634773. S2CID 2396763.
- ^ Callaway, Ewen (2017 yil 7-iyun). "Qadimgi Homo sapiens fotoalbomlari bizning turlarimiz tarixini qayta yozadi". Tabiat. doi:10.1038 / tabiat.2017.22114.
- ^ Tryon, Kristofer A.; Faith, Tyler (2013). "Sharqiy Afrikaning o'rta tosh asridagi o'zgaruvchanlik" (PDF). Hozirgi antropologiya. 54 (8): 234–54. doi:10.1086/673752. S2CID 14124486.
- ^ Mehta, Ankita (2018 yil 26-yanvar). "Isroilda topilgan 177 ming yillik jag 'suyagi toshqotganligi Afrikadan tashqarida topilgan eng qadimgi odam qoldiqlari". International Business Times. Olingan 28 yanvar 2018.
- ^ Zimmer, Karl (2019 yil 10-iyul). "Yunonistonda topilgan bosh suyagi suyagi insoniyat tarixini o'zgartirishi mumkin. G'orda topilgan suyak - bu Evropada topilgan eng qadimgi zamonaviy inson qoldiqlari. Odamlar Afrikani tark etishni bir paytlar o'ylamaganidan ancha ilgari boshlaganiga ishora qilmoqda". The New York Times. Olingan 11 iyul 2019.
- ^ Xodimlar (2019 yil 10-iyul). "'Afrikadan tashqaridagi eng qadimgi qoldiqlar inson migratsiyasi soatini tikladi ". Phys.org. Olingan 10 iyul 2019.
- ^ Harvati, Katerina; va boshq. (2019 yil 10-iyul). "Apidima g'orining qoldiqlari Evroosiyoda Homo sapiens haqida dastlabki dalillarni beradi". Tabiat. 571 (7766): 500–504. doi:10.1038 / s41586-019-1376-z. PMID 31292546. S2CID 195873640.
- ^ Xaynts, Tanja; Pala, Mariya; Gomes-Karballa, Alberto; Richards, Martin B.; Salas, Antonio (mart 2017). "Afrikalik odam mitoxondriyal DNK daraxtini yangilash: sud ekspertizasi va aholi genetikasi bilan bog'liqligi". Xalqaro sud ekspertizasi: Genetika. 27: 156–159. doi:10.1016 / j.fsigen.2016.12.016. PMID 28086175.
- ^ Poznik, G. D .; Xen, B. M.; Yee, M.-C .; Sliwerska, E .; Euskirchen, G. M .; Lin, A. A .; Snayder, M.; Kintana-Murchi, L.; Kidd, J. M .; Underhill, P. A .; Bustamante, C. D. (2013 yil 1-avgust). "Y xromosomalarini ketma-ketligi vaqt o'tishi bilan erkaklarning umumiy ajdodlari va ayollariga nisbatan farqni bartaraf etdi". Ilm-fan. 341 (6145): 562–565. Bibcode:2013 yil ... 341..562P. doi:10.1126 / science.1237619. PMC 4032117. PMID 23908239.
- ^ Karmin, Monika; Saag, Lauri; Visente, Mari; Sayres, Melissa A. Uilson; Mari, Mari; Talas, Ulvi Gerst; Rootsi, Siiri; Ilumäe, Anne-May; Magi, Reidik; Mit, Mario; Pagani, Luka; Puurand, Tarmo; Faltiskova, Zuzana; Klemente, Florian; Kardona, Aleksiya; Metspalu, Ene; Saxakyan, Ovannes; Yunusbayev, Bayazit; Xudjashov, Georgi; DeGiorgio, Maykl; Loogväli, Eva-Liis; Eyxstaedt, Kristina; Eelmets, Mikk; Chaubey, Gyaneshver; Tambets, Kristiina; Litvinov, Sergey; Mormina, Maru; Syu, Yali; Ayub, Qosim; va boshq. (2015 yil aprel). "Y xromosomalari xilma-xilligining so'nggi daragi madaniyatdagi global o'zgarishlarga to'g'ri keladi". Genom tadqiqotlari. 25 (4): 459–466. doi:10.1101 / gr.186684.114. PMC 4381518. PMID 25770088.
- ^ Klark, J. Desmond; Beyene, Yonas; VoldeGabriel, Giday; Xart, Uilyam K.; Renne, Pol R.; Gilbert, Genri; Defleur, Alban; Suva, Gen; Katoh, Shighero; Lyudvig, Kennet R.; Biskvit, Jan-Reno; Asfav, Berxan; Oq, Tim D. (2003 yil iyun). "Efiopiyaning O'rta Awash shahridan Pleistosen Homo sapiensning stratigrafik, xronologik va xulq-atvori". Tabiat. 423 (6941): 747–752. Bibcode:2003 yil natur.423..747C. doi:10.1038 / nature01670. PMID 12802333. S2CID 4312418.
- ^ Scerri, Eleanor (2017). "Afrika tarixining Oksford tadqiqotlari entsiklopediyasi". doi:10.1093 / acrefore / 9780190277734.013.137. ISBN 9780190277734. Iqtibos jurnali talab qiladi
| jurnal =(Yordam bering);| bob =mensimagan (Yordam bering) - ^ Henshilwood, C.S. va B. Dubreuil 2009. Artefaktlarni o'qish: Afrikaning janubidagi O'rta tosh asridan boshlab til yig'ish. R. Bota va C. Naytda (tahr.) Til beshigi. Oksford: Oksford universiteti matbuoti, 41-61 bet.
- ^ Bowler JM, Johnston H, Olley JM, Preskott JR, Roberts RG, Shawcross V, Spooner NA (2003). "Avstraliyaning Mungo ko'lida odamlarni ishg'ol qilish va iqlim o'zgarishi uchun yangi asrlar". Tabiat. 421 (6925): 837–40. Bibcode:2003 yil natur.421..837B. doi:10.1038 / tabiat01383. PMID 12594511. S2CID 4365526.
- ^ Richard E. Grin; Krause, J .; Briggs, A.W.; Maritsik, T .; Stenzel, U .; Kirxer, M .; Patterson, N .; Li, X.; va boshq. (2010). "Neandertal genomining navbatdagi loyihasi". Ilm-fan. 328 (5979): 710–22. Bibcode:2010Sci ... 328..710G. doi:10.1126 / science.1188021. PMC 5100745. PMID 20448178.
- ^ Rincon, Pol (2010-05-06). "Neandertal genlari bizda omon qoladi'". BBC yangiliklari. BBC. Olingan 2010-05-07.
- ^ Sankararaman, Sriram; Mallik, svopen; Patterson, Nik; Reyx, Devid (2016). "Hozirgi odamlarda Denisovan va neandertal ajdodlarining qo'shma manzarasi". Hozirgi biologiya. 26 (9): 1241–1247. doi:10.1016 / j.cub.2016.03.037. PMC 4864120. PMID 27032491.
- ^ Callaway, Ewen (2012 yil 26-iyul). "Ovchi-yig'uvchi genom genetik xilma-xillikni keltirib chiqaradi". Tabiat. doi:10.1038 / tabiat.2012.11076. S2CID 87081207.
- ^ Laxans, Jozef; Vernot, Benjamin; Elbers, Klara S.; Ferverda, Bart; Froment, Alen; Bodo, Jan-Mari; Lema, Godfri; Fu, Venqing; Nyambo, Tomas B.; Rebbek, Timoti R.; Chjan, Kun; Akey, Joshua M.; Tishkoff, Sara A. (2012 yil avgust). "Evolyutsion tarix va turli xil Afrikalik ovchi-yig'uvchilarning yuqori qamrovli butun genomli ketma-ketliklaridan moslashuvi". Hujayra. 150 (3): 457–469. doi:10.1016 / j.cell.2012.07.009. PMC 3426505. PMID 22840920.
- ^ Xu, Duo; Pavlidis, Pavlos; Taskent, Recep O'zgur; Alachiotis, Nikolaos; Flanagan, Kolin; DeGiorgio, Maykl; Blexman, Ran; Rul, Stefan; Gokcumen, Omer (2017 yil oktyabr). "Afrikadagi arxaik homininning tajovuzi funktsional tuprik MUC7 genetik o'zgarishiga yordam beradi". Molekulyar biologiya va evolyutsiya. 34 (10): 2704–2715. doi:10.1093 / molbev / msx206. PMC 5850612. PMID 28957509.
- ^ Mondal, Mayux; Bertranpetit, Xaume; Lao, Oskar (2019 yil 16-yanvar). "Chuqur o'rganish bilan Bayesning taxminiy hisob-kitoblari Osiyo va Okeaniyada uchinchi arxaik introressiyani qo'llab-quvvatlaydi". Tabiat aloqalari. 10 (1): 246. Bibcode:2019NatCo..10..246M. doi:10.1038 / s41467-018-08089-7. PMC 6335398. PMID 30651539.
- ^ Klayn, Richard (1995). "Anatomiya, xulq-atvor va zamonaviy inson kelib chiqishi". World Prehistory jurnali. 9 (2): 167–98. doi:10.1007 / bf02221838. S2CID 10402296.
- ^ Sutikna, Tomas; Tocheri, Metyu V.; Morvud, Maykl J.; Saptomo, E. Vaxyu; Jatmiko; Qo'rquv, Rokus tufayli; Vasisto, Shri; Westaway, Kira E.; Obert, Maksim; Li, Bo; Chjao, Tszian-xin; Stori, Maykl; Alloway, Brent V.; Morley, Mayk V.; Meijer, Hanneke JM.; van den Berg, Gerrit D.; Grün, Rayner; Dosseto, Entoni; Brumm, Odam; Jungers, Uilyam L.; Roberts, Richard G. (2016 yil 30 mart). "Indoneziyadagi Liang Buada homo floresiensis uchun qayta ko'rib chiqilgan stratigrafiya va xronologiya". Tabiat. 532 (7599): 366–69. Bibcode:2016Natur.532..366S. doi:10.1038 / tabiat17179. PMID 27027286. S2CID 4469009.
- ^ Belesal, Sandra; Santos, AM; McEvoy, B .; Alves, men.; Martinho, C .; Kemeron, E .; Shriver, MD; Parra, EJ .; Rocha, J. (2012). "Evropaliklarda pigmentatsiyani yoritish vaqti". Molekulyar biologiya va evolyutsiya. 30 (1): 24–35. doi:10.1093 / molbev / mss207. PMC 3525146. PMID 22923467.
- ^ Fumagalli, M.; Moltke, I .; Grarup, N .; Rakimo, F.; Byerregaard, P .; Yorgensen, M. E.; Korneliussen, T. S.; Gerbault, P .; Skotte, L .; Linneberg, A .; Kristensen, C .; Brandslund, I .; Yorgensen, T .; Xerta-Sanches, E .; Shmidt, E. B.; Pedersen, O .; Xansen, T .; Albrechtsen, A .; Nilsen, R. (17 sentyabr 2015). "Grenlandiyalik Inuit ovqatlanish va iqlimga moslashishning genetik imzosini namoyish etadi". Ilm-fan. 349 (6254): 1343–1347. Bibcode:2015 yil ... 349.1343F. doi:10.1126 / science.aab2319. hdl:10044/1/43212. PMID 26383953. S2CID 546365.
- ^ Peng, Yi; Shi, Xong; Qi, Xue-bin; Syao, Chun-tsie; Chjun, Xua; Ma, Run-lin Z; Su, Bing (2010). "Sharqiy Osiyo populyatsiyasida ADH1B Arg47His polimorfizmi va tarixda guruchni xonakilashtirishning kengayishi". BMC evolyutsion biologiyasi. 10 (1): 15. doi:10.1186/1471-2148-10-15. PMC 2823730. PMID 20089146.
- ^ Segurel, Laure; Bon, Céline (2017 yil 31-avgust). "Odamlarda laktaza qat'iyligi evolyutsiyasi to'g'risida". Genomika va inson genetikasining yillik sharhi. 18 (1): 297–319. doi:10.1146 / annurev-genom-091416-035340. PMID 28426286.
- ^ Ingram, Ketrin J. E .; Mulkrey, Sharlotta A.; Itan, Yuval; Tomas, Mark G.; Qaldirg'och, Dallas M. (2008 yil 26-noyabr). "Laktoza hazm bo'lishi va laktaza doimiyligining evolyutsion genetikasi". Inson genetikasi. 124 (6): 579–591. doi:10.1007 / s00439-008-0593-6. PMID 19034520. S2CID 3329285.
Tashqi havolalar
- Paleoslar
- Hominid yilnomasi
- Berkli Evolyutsiyasi
- History of Animal Evolution
- Hayot daraxti veb-loyihasi – explore complete phylogenetic tree interactively
- Inson xronologiyasi (interaktiv) – Smithsonian, Milliy tabiiy tarix muzeyi (Avgust 2016).
| Sub-mavzular | |
|---|---|
| Genetika tarixi mintaqalar bo'yicha | |
| Populyatsiya genetikasi guruh bo'yicha |
|
| |
| Taksonomiya (Gomininlar ) |
| ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ajdodlar |
| ||||||||||||||||||||||||||
| Modellar |
| ||||||||||||||||||||||||||
| Vaqt jadvallari | |||||||||||||||||||||||||||
| Boshqalar |
| ||||||||||||||||||||||||||
| |||||||||||||||||||||||||||

{kind=link}